Meiosis I
Homologous chromosomes separate in meiosis I
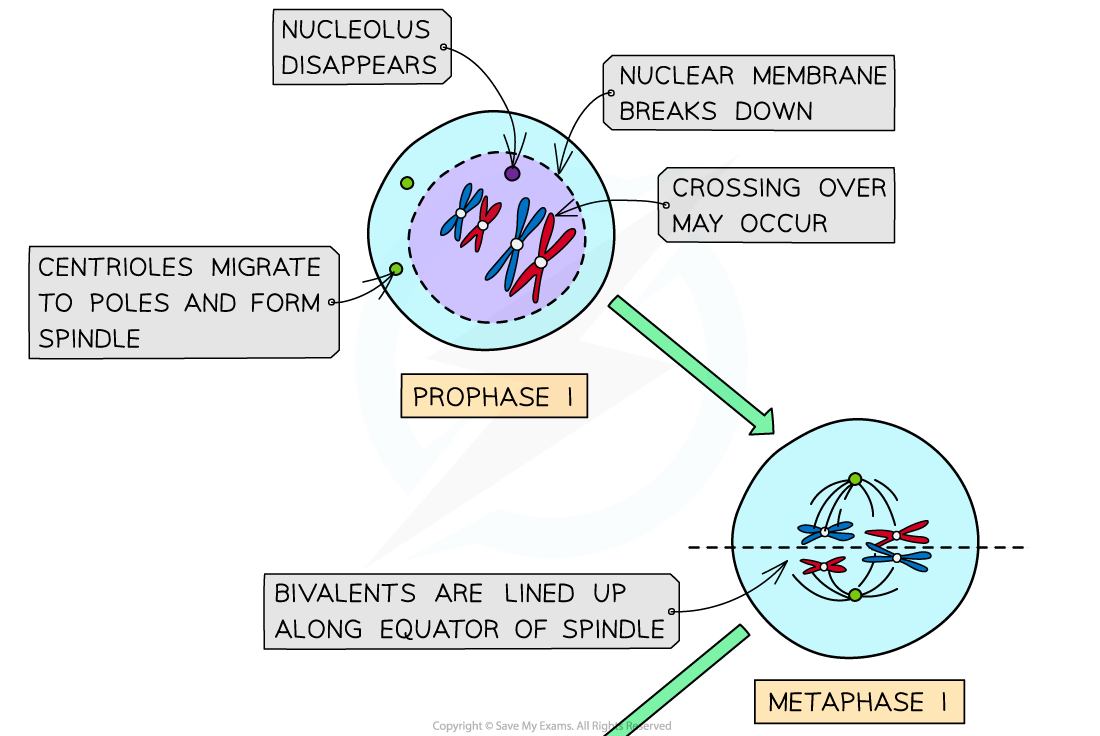
Prophase I
- DNA has already replicated and condenses and becomes visible as chromosomes
- Each chromosome consists of two sister chromatids joined together by a centromere
- The chromosomes are arranged side-by-side in homologous pairs
- A pair of homologous chromosomes is called a bivalent
- As the homologous chromosomes are very close together the crossing over of non-sister chromatids may occur. The point at which the crossing over occurs is called the chiasma (chiasmata; plural)
- In this stage centrioles migrate to opposite poles and the spindle is formed
- The nuclear envelope breaks down and the nucleolus disintegrates
Metaphase I
- The bivalents line up along the equator of the spindle, with the spindle fibres attached to the centromeres
- The bivalents line up by independent assortment (random orientation)
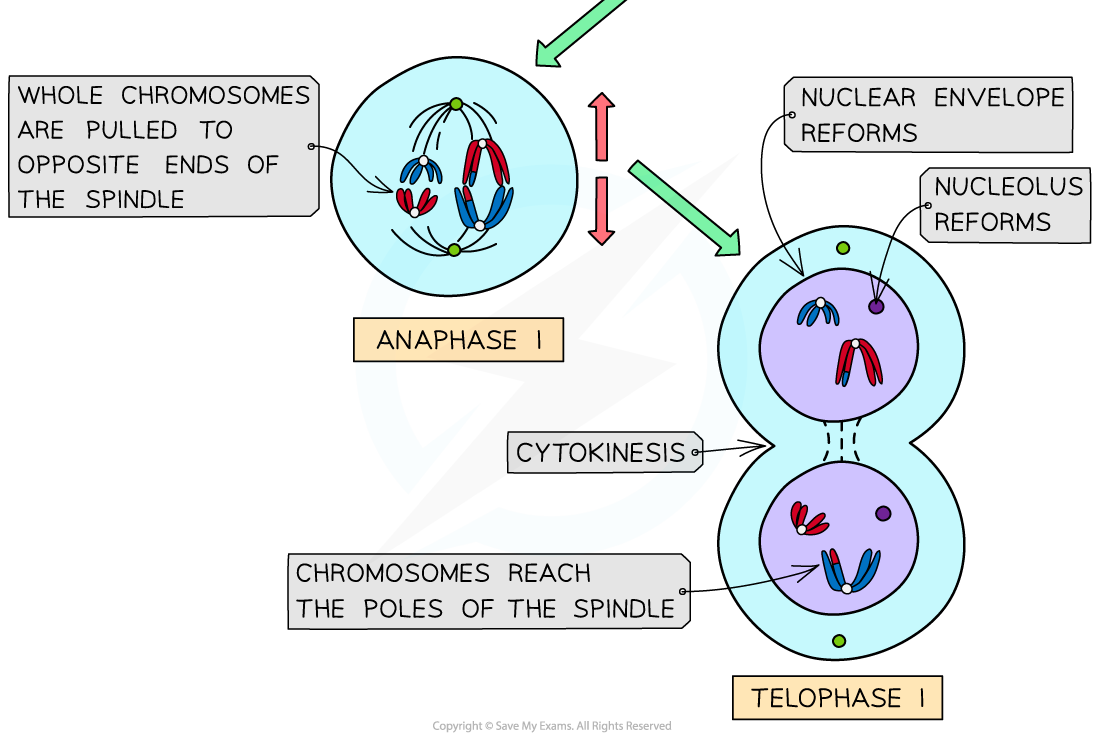
Anaphase I
- The homologous pairs of chromosomes are separated as microtubules pull whole chromosomes to opposite ends of the spindle
- The centromeres do not split
Telophase I
- The chromosomes arrive at opposite poles
- Spindle fibres start to break down
- Nuclear envelopes form around the two groups of chromosomes and nucleoli reform
- Some plant cells go straight into meiosis II without reformation of the nucleus in telophase I
Meiosis I is reduction division
- Meiosis I is referred to as reduction division because homologous chromosomes separate and move to opposite poles of the cell.
- Therefore, the number of chromosomes per cell is reduced by a factor of 2
The different stages of meiosis I in an animal cell
Exam Tip
Understanding the difference between chromosomes and chromatids can be difficult. We count chromosomes by the number of centromeres present. So when the 46 chromosomes duplicate during interphase and the amount of DNA in the cell doubles there are still only 46 chromosomes present because there are still only 46 centromeres present. However, there are now 92 chromatids, which are strands of replicated chromosomes.
Independent Assortment
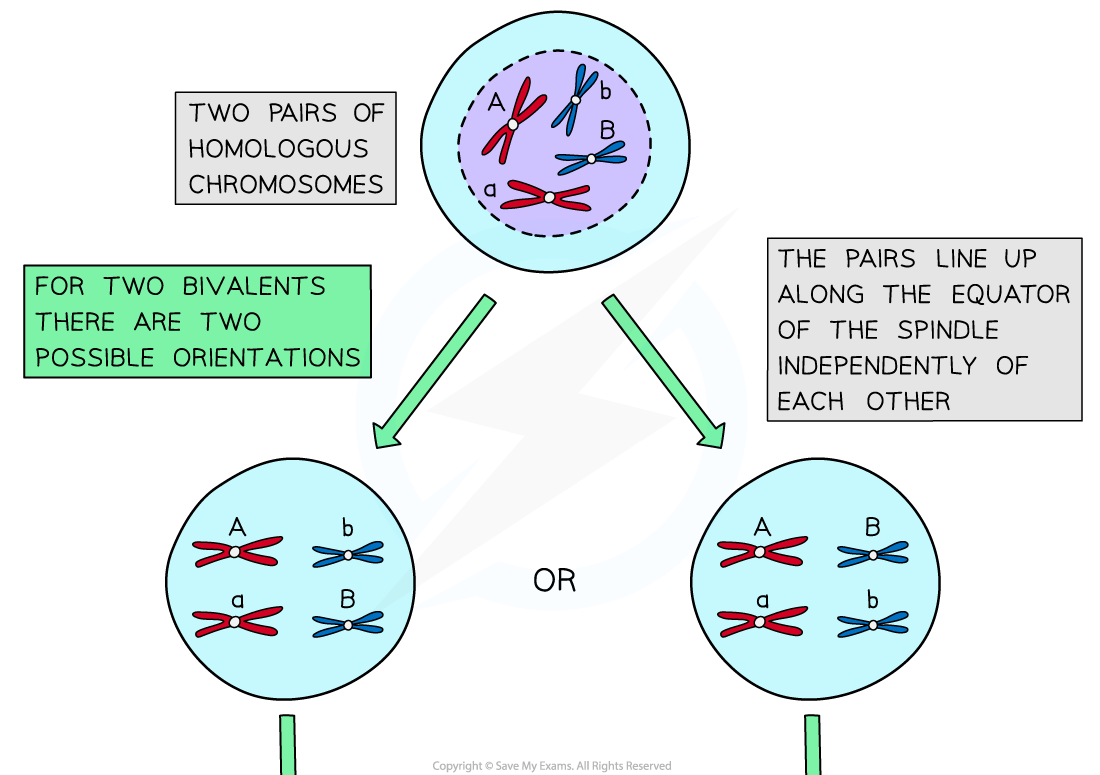
- During metaphase I, an event occurs that is an important source of genetic variation in the gametes formed by meiosis
- Independent assortment is the production of different combinations of alleles in gamete cells due to the random alignment of homologous pairs along the equator of the spindle during metaphase I
- This random alignment is sometimes referred to as random orientation
- In prophase I homologous chromosomes pair up and in metaphase I they are pulled towards the equator of the spindle
- Each pair can be arranged with either chromosome on top, this is completely random
- The orientation of one homologous pair is independent / unaffected by the orientation of any other pair
- The homologous chromosomes are then separated and pulled apart to different poles during anaphase
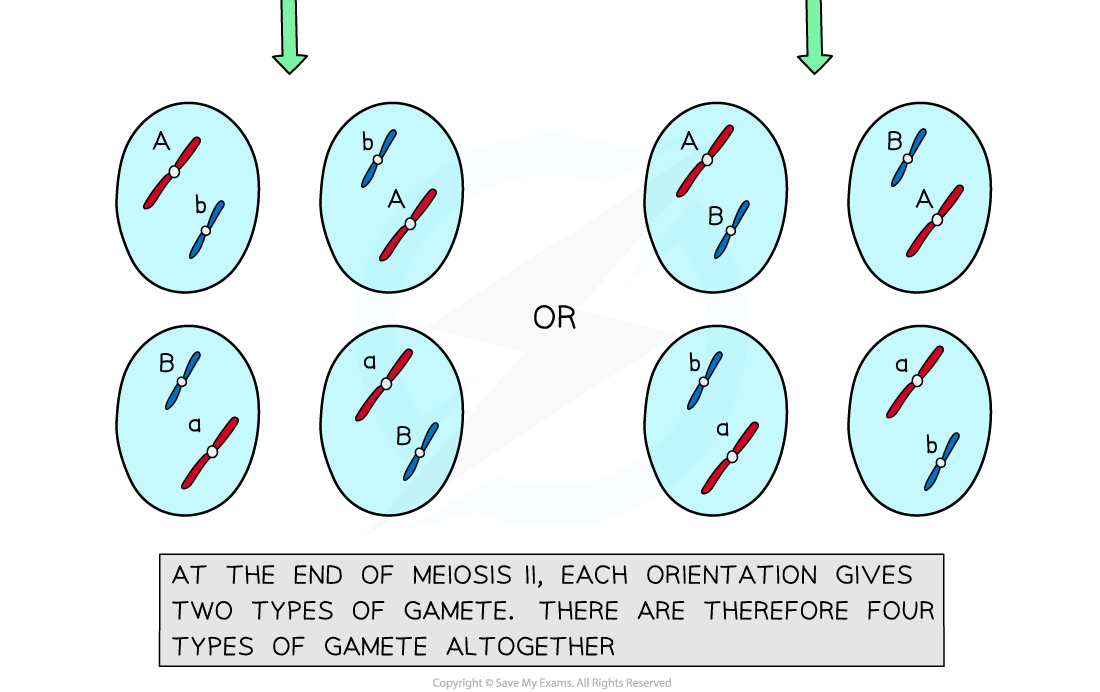
- The combination of alleles that end up in each daughter cell depends on how the pairs of homologous chromosomes were lined up
- To work out the number of different possible chromosome combinations the formula 2n can be used, where n corresponds to the number of chromosomes in a haploid cell
- For humans this is 223 which calculates as 8 388 608 different combinations
- This may seem like a lot of combinations, but by contrast, crossing over introduces an almost infinite amount of variation
Independent assortment of homologous chromosomes leading to different genetic combinations in daughter cells
Explaining Deviations from Mendelian Ratios
NOS: Careful observation and record keeping turned up anomalous data that Mendel’s law of independent assortment could not account for
- Gregor Mendel (of pea plant fame) devised the law of independent assortment in 1866
- In this, he states that characteristics are inherited completely independently of others
- This was refined by later scientists to state that the allele that gets sorted into a particular gamete cell is not influenced by the allele received for another gene
- In many cases, this holds true, and can be observed in Mendelian ratios of offspring in certain crossing experiments
- Mendel observed a 9:3:3:1 ratio in many dihybrid crosses of sweet pea plants
- As a result, Mendel's findings were not challenged until the early 20th century, when Bateson and Punnett found seemingly anomalous ratios of offspring
- For which they could offer no explanation
- These became named as non-Mendelian ratios because they did not follow the pattern as predicted by Mendel
Bateson and Punnetts' experiment (1905)
- Also working with sweet peas, two pairs of alleles were identified (Dominant alleles in capital letters)
- Flower colour:
- P = purple, p = red
- The shape of pollen grains:
- R = long, r = round
- Flower colour:
- Their first cross involved pure-bred purple-flowered plants with long pollen grains (PPRR) with pure-bred red-flowered plants with round pollen grains (pprr)
- As expected, this cross resulted in 100% purple-flowered plants with long pollen grains, with the double-heterozygous genotype PpRr in the F1 generation
- However, when individuals from the F1 generation were crossed with each other, Bateson and Punnett would have expected a 9:3:3:1 mix of phenotypes, in line with Mendel's law of independent assortment
Modelling the expected ratios using a Punnet square
Dihybrid Cross Punnett Square Table

- Expected phenotype ratios for the second generation:
- 9 purple flowers, long pollen grains
- 3 purple flowers, round pollen grains
- 3 red flowers, long pollen grains
- 1 red flowers, round pollen grains
Actual Results from Bateson and Punnetts' Experiment
- In the F2 generation of offspring, there were...
- Very many of the grandparent phenotype (purple-flowered, long pollen grains) produced
- Many of the grandparent phenotype (red-flowered, round pollen grains) produced
- Very few (but not zero) other phenotypes (purple/round or red/long) produced
- This appeared as an anomaly to Bateson and Punnett, though they did not find an explanation
Thomas Hunt Morgan developed the notion of linked genes to account for the anomalies
- Later in the 20th century (approx. 1910-1940), an American biologist, Thomas Hunt Morgan, put forward an explanation for the results previously observed by Bateson and Punnett
- From his findings, he devised the theory of linkage
- Morgan worked on fruit flies (Drosophila melanogaster) thanks to their ability to reproduce quickly, in large numbers, in a small physical space and with observable heritable characteristics like eye colour and wing shape
- In fact, Morgan bred them selectively for many years to develop a range of phenotypes by natural mutation
- Morgan's work identified sex-linked characteristics from genes carried on the X or Y chromosomes that determine gender
- This work led Morgan to develop the idea of autosomal linkage
- This is where unexpected patterns of inheritance are caused by separate alleles being inherited together, from the same chromosome (an autosome)
- Morgan then elaborated his work by developing the theory of crossing over
- As a way of accounting for unexpected (recombinant) genotypes
Exam Tip
Several sources of genetic variation have been outlined above. It is also worth remembering that genetic variation can occur on an even smaller scale than chromosomes. Mutations can occur within genes. A random mutation that takes place during DNA replication can lead to the production of new alleles and increased genetic variation.
